L’homéostasie tissulaire et la régénération nécessitent un équilibre précis entre la quiescence et l'activation des cellules souches adultes. L'intérêt principal de notre laboratoire est de comprendre les mécanismes moléculaires et cellulaires qui régulent cet équilibre. Nous utilisons la peau en tant que système modèle pour étudier le comportement des cellules souches adultes et disséquer les mécanismes sous-jacents régulant la fonction des cellules souches.
La peau est le plus grand organe du corps. Il est composé de deux grands compartiments: un compartiment externe épithélial (épiderme) et un compartiment mésenchymateuses interne (derme). Le compartiment épithélial est composé de l'épiderme et de ses annexes: les follicules pileux et les glandes sébacées. Le compartiment mésenchymateux est en grande partie composé par des cellules fibroblastes dispersées dans la matrice extracellulaire.
L'épiderme est en équilibre dynamique et se renouvelle sans cesse tout au long de la vie. Les cellules souches épithéliales de la peau peuvent s'auto-renouveler et se différencier afin de fournir la source illimitée de cellules nécessaires à la morphogenèse à long terme des tissus, l'homéostasie, et la réparation des blessures. Les cellules souches épithéliales de la peau comprennent des cellules de l'épiderme de la tige, qui sont localisées dans la couche interfolliculaire épidermique de la base, et les cellules souches du follicule pileux, qui résident dans une région de la gaine externe du bulbe du follicule pileux (ORS) connu en tant que le renflement. La morphogénèse du follicule pileux est particulièrement intéressante en ce qu'elle implique une série temporelle d’interactions épithélio-mésenchymateuses. L'épiderme embryonnaire commence comme une couche simple épithéliale, et lorsque le mésenchyme remplit la peau, le développement du derme transmet les premiers signaux à l'épiderme pour faire un appendice. En réponse, l'épiderme envoie un signal aux cellules dermiques de se condenser et former la papille dermique (DP) qui deviendra un composant essentiel et permanent de chaque follicule pileux. Un autre signal est ensuite libéré de la DP afin de promouvoir la prolifération et la différenciation du follicule pileux.
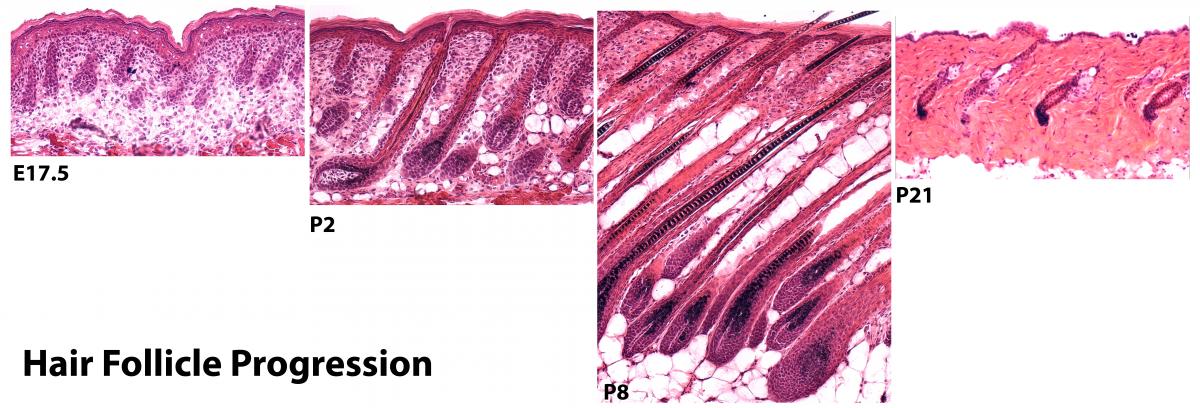
A mi-chemin à travers la maturation du follicule pileux en développement, un groupe de cellules au cycle de division cellulaire rare apparaît dans la partie supérieure du follicule pileux, juste en dessous de l'endroit où la glande sébacée émergera. Ces cellules à cycle lent résidant dans la région du bulge sont appelées cellules souches du follicule pileux (HFSCs). Pendant la phase de croissance des poils en régénération normale, HFSCs soit restent dans la région de renflement ou donnent lieu à une descendance qui sortira le long des SRO. Les cellules GEB produisent alors des cellules à prolifération rapide et amplification transitoire (TAC) appelées cellules de la matrice. Tout au long de la phase du cycle de croissance pilaire adulte, le DP réside à la base même du follicule, où il agit comme un centre de signalisation pour l’interaction avec les cellules de la matrice afin de les stimuler pour proliférer, se différencier et produire le poil et son canal, ou gaine interne (IRS).
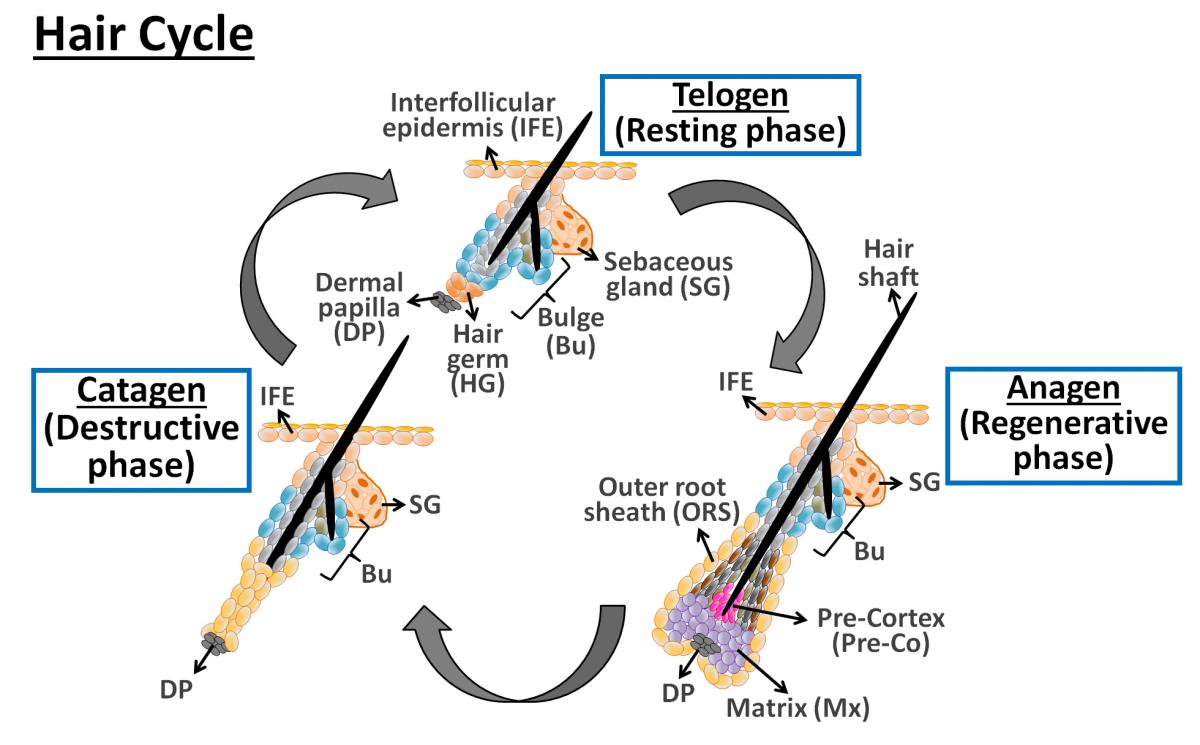
Les cellules souches du follicule du poil dans la zone de renflement sont maintenues dans un état de repos au cours de la phase de repos du cycle pileux. Au cours de la croissance du poil, elles sont activées et remplacent les cellules dans le follicule pileux inférieur. En outre, ces cellules souches peuvent se déplacer vers le haut et se différencier en épiderme et glandes sébacées et réparer, au moins de façon transitoire, l'épithélium lésé pendant la cicatrisation des plaies. Qu'est-ce qui maintient les cellules souches dans leur niche pendant les périodes de repos et qu’est-ce qui les incite (ou leur descendance immédiate) à régénérer le follicule pendant la croissance des poils ou la réparation de l'épiderme après blessure?
Une étude de profil transcriptionnel des cellules souches purifiées dans le renflement du follicule des poils adultes a fourni une signature moléculaire qui offre des perspectives sur les mécanismes possibles. Couplées avec des études du profil génétique, des études fonctionnelles ultérieures révèlent plusieurs voies importantes pour le maintien des cellules souches soit dans un état de repos ou activé. L'étude des mécanismes de la façon dont les cellules souches sont régulées est importante pour la médecine régénérative et aussi pour le traitement de pathologies telles que le cancer. Dans notre laboratoire, nous utilisons la souris comme un organisme modèle pour répondre à cette question biologique importante. L'intérêt principal de notre laboratoire est de comprendre comment une voie de signalisation s’intègre à une autre et comment leurs régulateurs en aval interagissent dans les cellules souches et / ou avec les cellules voisines aux niveaux moléculaires et cellulaires. Nos études nous aideront à mieux comprendre comment les cellules souches fonctionnent dans des organes adultes définis, et améliorer notre façon d'utiliser les cellules souches pour traiter certaines maladies telles que le cancer.
The epidermis is in dynamic equilibrium and constantly renewed throughout life. Skin epithelial stem cells that can self-renew and differentiate provide the unlimited source of cells required for long-term tissue morphogenesis, homeostasis, and injury repair. Skin epithelial stem cells encompass epidermal stem cells, which are localized in interfollicular epidermal basal layer, and hair follicle stem cells, which reside in a region of the hair follicle outer root sheath known as the bulge.
Hair follicle stem cells, residing at the bulge region, are maintained in a quiescent state during the resting phase of hair cycle. During hair growth, they then get activated and replace the cells in the lower hair follicle, or move upward and differentiate into the epidermis and sebaceous gland during wound healing. Tissue homeostasis and regeneration require a precise balance between quiescence and activation of adult stem cells. What keeps stem cells in their bulge niche during times of quiescence and what prompts them or their immediate progeny to regenerate the follicle during hair growth or repair the epidermis upon wounding?
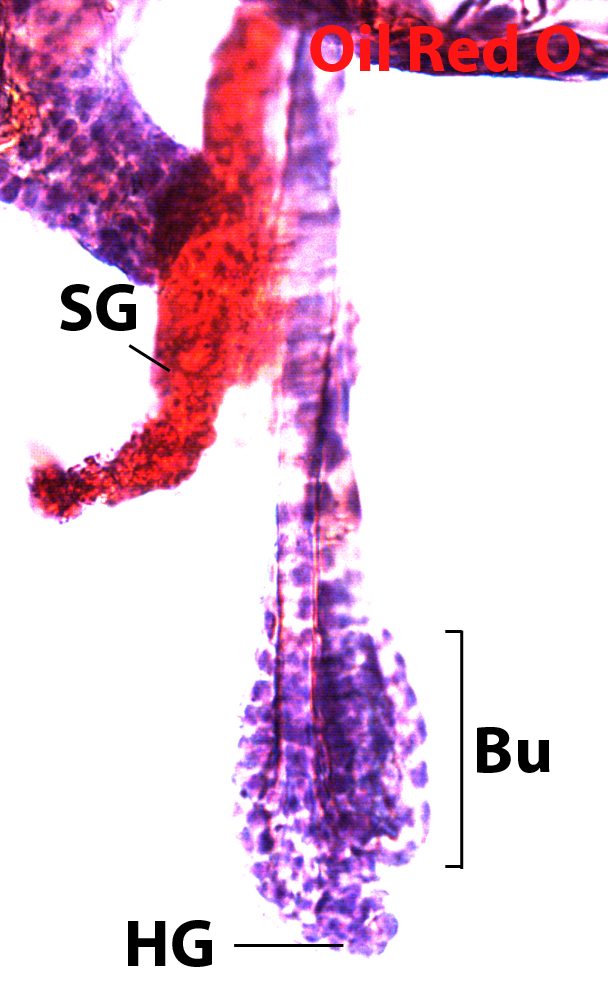
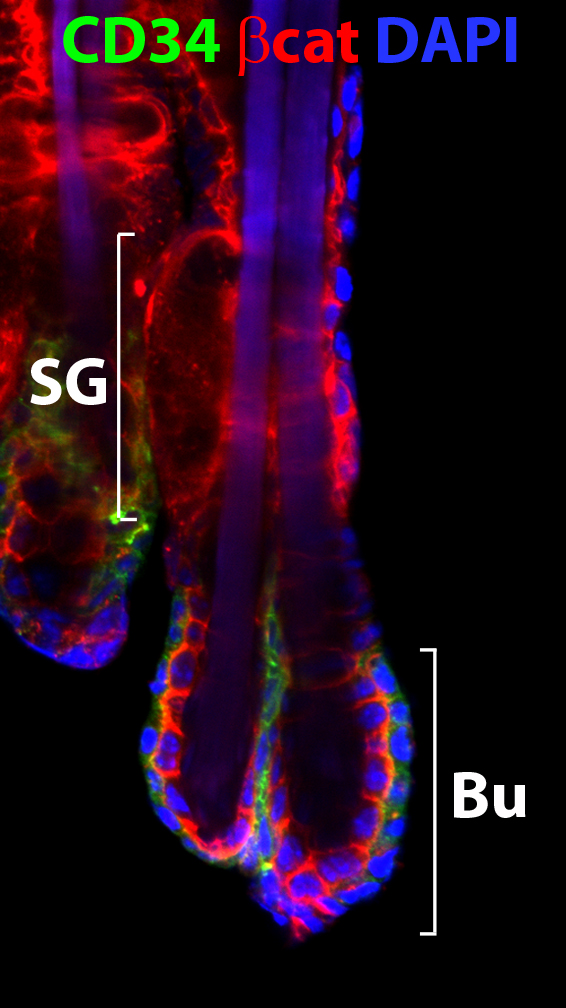

The behavior of stem cell is fine-tuned by the microenvironmental cues and their responsive intracellular signaling regulation. Several signaling pathways, including BMP, Wnt and TGFβ signaling pathways, are important for maintaining the stem cells in either a quiescent or activated state. Studying the mechanisms of how stem cells are regulated is important for regenerative medicine and also for treatment of pathological conditions such as cancer. In our lab, we use mouse as a model organism to address this important biological question. Coupling with multiple technologies, such as transgenic mouse model, fluorescent activated cell sorting (FACS), chromatin-immunoprecipitation sequencing (ChIP-seq), primary cell culture, and proteomic approaches, we want to better understand how one signaling pathway integrates to another and how their downstream regulators cross-talk to each other in stem cells and/or with neighboring cells in molecular and cellular levels. Our studies will help us appreciate how stem cells function in the defined adult organs, and how to use stem cells to treat diseases and cancer.
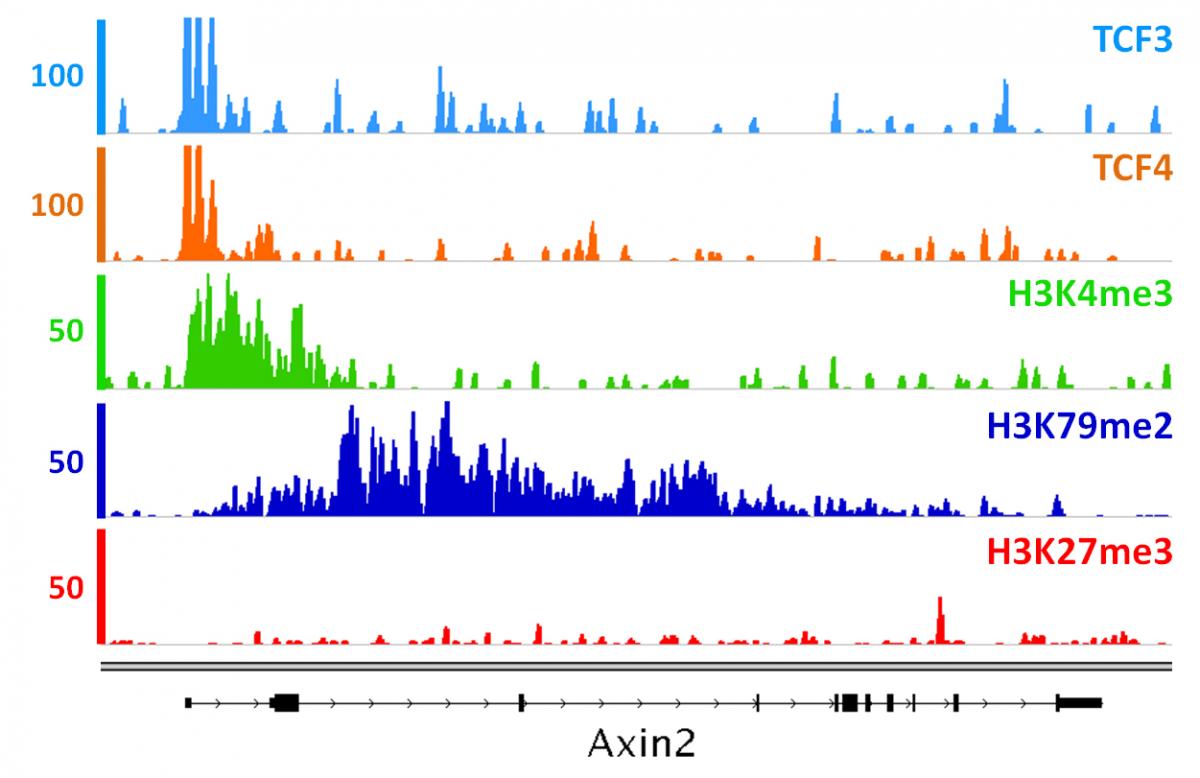
Jossin Y, Lee M, Klezovitch O, Kon E, Cossard A, Lien WH, Fernandez TE, Cooper JA, Vasioukhin V.
Dev Cell. 2017; 41(5):481-95.e5.
Li P, Silvis MR, Honaker Y, Lien WH, Arron ST, Vasioukhin V.
Genes Dev. 2016; 30(7):798-811.
Wang W, Jossin Y, Chai G, Lien WH, Tissir F, Goffinet AM.
Nat Commun. 2016; 7:10936.
Lien WH, Fuchs E.
Genes Dev. 2014; 28(14):1517-32.
Rockowitz S, Lien WH, Pedrosa E, Wei G, Lin M, Zhao K, Lachman HM, Fuchs E, Zheng D.
PLoS Comput Biol. 2014; 10(6):e1003671.
Lien WH, Polak L, Lin M, Lay K, Zheng D, Fuchs E.
Nat Cell Biol. 2014; 16(2):179-90.
Keyes BE, Segal JP, Heller E, Lien WH, Chang CY, Guo X, Oristian DS, Zheng D, Fuchs E.
Proc Natl Acad Sci U S A. 2013; 110(51):E4950-9.
Beronja S, Janki P, Heller E, Lien WH, Keyes BE, Oshimori N, Fuchs E.
Nature. 2013; 501(7466):185-90.
Bock C, Beerman I, Lien WH, Smith ZD, Gu H, Boyle P, Gnirke A, Fuchs E, Rossi DJ, Meissner A.
Mol Cell. 2012; 47(4):633-47.
Lien WH, Guo X, Polak L, Lawton LN, Young RA, Zheng D, Fuchs E.
Cell Stem Cell. 2011; 9(3):219-32.
Silvis MR, Kreger BT, Lien WH, Klezovitch O, Rudakova GM, Camargo FD, Lantz DM, Seykora JT, Vasioukhin V.
Sci Signal. 2011; 4(174):ra33.
Ezhkova E, Lien WH, Stokes N, Pasolli HA, Silva JM, Fuchs E.
Genes Dev. 2011; 25(5):485-98.
Mendrysa SM, Akagi K, Roayaei J, Lien WH, Copeland NG, Jenkins NA, Eisenman RN.
Lien WH, Gelfand VI, Vasioukhin V.
J Cell Biol. 2008; 183(6):989-97.
Lien WH, Stepniak E, Vasioukhin V.
Proc Natl Acad Sci U S A. 2008; 105(40):15225-6.
Lien WH, Klezovitch O, Null M, Vasioukhin V.
J Cell Sci. 2008; 121(Pt 9):1357-62.
Lien WH, Klezovitch O, Vasioukhin V.
Curr Opin Cell Biol. 2006; 18(5):499-506. Review.
Lien WH, Klezovitch O, Fernandez TE, Delrow J, Vasioukhin V.
Science. 2006; 311(5767):1609-12.

 Telecharger
Telecharger
INTERACTIONS DE VOIES DE SIGNALISATION DANS LES CELLULES SOUCHES DE LA PEAU
